¿Que debo saber sobre la relación de la Cafeína con las enfermedades de mis pacientes?

Figura
1 Estructura molecular de la cafeína. La cafeína
es parte de un
grupo de compuestos llamados
trimetilxantinas, que incluyen
teobromina y teofilina
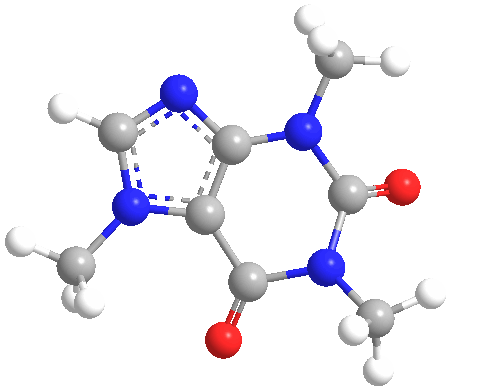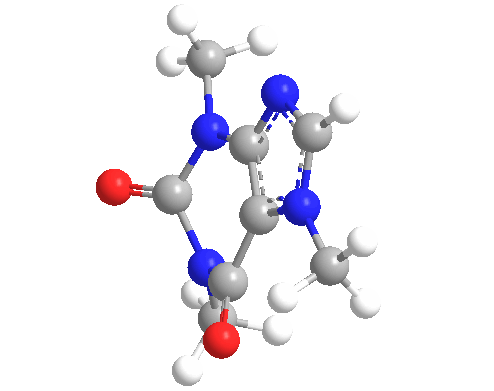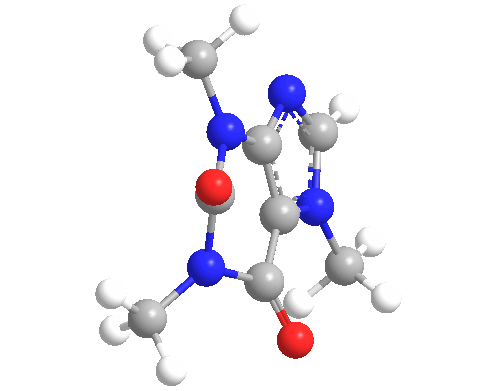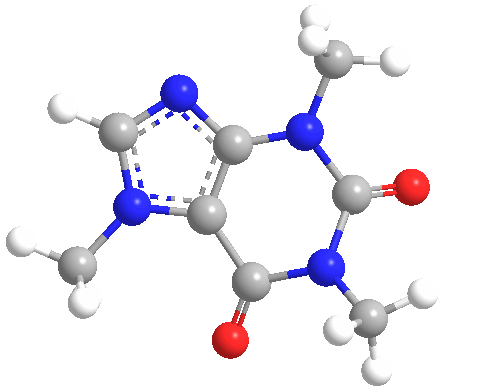
Figura
1 Estructura molecular de la cafeína. La cafeína
es parte de un
grupo de compuestos llamados
trimetilxantinas, que incluyen
teobromina y teofilina
Fuentes y propiedades químicas de la cafeína
La cafeína es el agente psicoactivo más consumido en el mundo. Se trata de una sustancia natural, que puede encontrarse en más de 60 especies de plantas, incluyendo té, nueces de cola, granos de café, hojas de mate, plantas de guaraná y nueces de cacao. Descubierta en 1819 por Friedrich Ferdinand Runge, que le dio nombre (Kaffein). A finales del siglo XIX Hermann Emil Fischer determinó la estructura de la cafeína y la sintetizó por primera vez. También se la conoce como guaranina, teína o mateína, según la planta de la que procede. La fuente principal de cafeína en la dieta humana es el consumo de café; pero también existen fármacos, en cuya composición entra la cafeína (Tablas 1 y 2 respectivamente).
 |
Tabla 1: Concentración de cafeína en diferentes presentaciones de bebidas e infusiones
La Organización de Consumidores y Usuarios dispone, en su sitio web de una calculadora de cafeína, que permite calcular el contenido de esta sustancia en diferentes bebidas y en fármacos (https://www.ocu.org/alimentacion/cafe/calculadora/cafeina) (1).
 |
Tabla 2: Contenido de cafeína sola o asociada a otros fármacos
La cafeína (C8H10N4O2) es una metilxantina (1,3,7-trimetilxantina o 3,7-dihidro-1,3,7-trimetil-1H-purina-2,6-diona) (Figura 1), que antagoniza los receptores de la adenosina (A1, A2A, A2B y A3), de forma no selectiva, en el tejido nervioso. Sus metabolitos incluyen la paraxantina, teobromina y teofilina (Figura 2), que tienen estructuras similares.
 |
| Figura 2. Metabolitos de la cafeína. FD. CC BY-SA 4.0. Wikimedia Commons |
Farmacocinética
Por vía digestiva. La cafeína se absorbe en estómago e intestino delgado y alcanza un pico aproximadamente a los 45 minutos tras la ingesta, independientemente de la fuente de la que proceda en la dieta. Los efectos subjetivos (menor somnolencia y mayor activación) suelen aparecer a los 10 minutos del consumo y son mas marcados en hombres, sin diferencias en cuanto a las horas del día (2).
Un estudio que comparó la magnitud y rapidez de los niveles máximos de cafeína y los efectos subjetivos entre el café y la cola, observó que no existía un efecto del vehículo de administración (infusión de café, bebida de cola o cápsulas de cafeína) o interacción del mismo con el fármaco, sobre la magnitud del efecto máximo o el tiempo hasta la aparición de dicho efecto. Aunque pueden observarse diferencias que pueden deberse a diferencias en la dosis, la hora del día, el edulcorante agregado o al entorno ambiental (3).
Los niveles de cafeína en saliva reflejan aceptablemente la concentración de cafeína en plasma libre y pueden usarse para estimar los parámetros farmacocinéticos del fármaco. En general, la relación de concentración de saliva/plasma es de 0,74 ± 0.08, con cierta dependencia temporal de la proporción, que es mas elevada inicialmente (incluso después de la administración intravenosa) y proporciones más bajas a intervalos de tiempo más largos después de la dosis. La eliminación urinaria de cafeína fue baja e independiente de la dosis: el 1,83% de la dosis se eliminó sin cambios. Presenta una farmacocinética lineal en un rango de dosis investigado de administración oral de 50-750 mg. e intravenosa de 300-750 mg.. Se absorbe rápidamente en todas las dosis y su tasa de eliminación aparente de primer orden disminuye linealmente con la dosis (0,163 ± 0,081 h−1 para 50 mg y 0,098 ± 0,027 h−1 para 750 mg.). En cuanto al aclaramiento total es de 0,98 ± 0,38 ml/min/kg. y no parece verse afectado por la dosis. Con una relación lineal entre el área bajo la concentración plasmática, la curva de tiempo y la dosis y la concentración plasmática normalizada por dosis (4).
Metabolismo de la cafeína
La cafeína se metaboliza casi por completo en hígado por la vía del Citocromo P450 (CYP) y solo se excreta menos del 3% sin cambios en la orina. La vía metabólica mas importante en humanos (70-80%) es la desmetilación de N-3 a paraxantina, también conocida como 1,7-dimetilxantina o 17X (en ratas la principal vía metabólica es la 8-hidroxilación). Tanto en humanos como en ratas esta reacción es llevada a cabo por la isoforma CYP1A2 en el hígado. Los experimentos con microsomas hepáticos humanos estiman que la 1-N-desmetilación a teobromina representa aproximadamente del 7 al 8% del metabolismo de la cafeína, con 7-N-desmetilación a teofilina también alrededor del 7 al 8%. El 15% restante de cafeína sufre hidroxilación C-8 para formar ácido 1,3,7-trimetilúrico (5, 6).
 |
Figura 3: Figura X. Vías metabólicas de la cafeina. (Abreviaturas: 1U, acido 1-metilurico; 1X, 1-metilxantina;17U, acido 1,7-dimetilurico; 17X, 1,7-dimetilxanthina; AFMU, 5-acetilamino-6-formilamino-3- metiluracilo; NAT-2, N-acetil-transferasa-2; XO, xantina oxidasa) |
La isoforma CYP1A2 es responsable de más del 95% del metabolismo primario de la cafeína. Por lo tanto, la cafeína se utiliza como fármaco sonda para la actividad de CYP1A2 y las proporciones relativas de metabolitos urinarios se utilizan como indicador del flujo a través de diferentes partes de la vía (BEGAS). Además de la paraxantina, los principales metabolitos de la cafeína en la orina son 1-metilxantina (1X), ácido 1-metilúrico (1U), 5-acetilamino-6-formilamino-3-metiluracilo (AFMU), ácido 1,7-dimetilúrico ( 17U) (Figura 4). Estos están formados por el metabolismo secundario de la paraxantina por CYP1A2, CYP2A6, NAT2 y XDH (también conocida como xantina oxidasa o XO) (BEGAS). Los estudios in vitro en líneas celulares muestran la participación de CYP2E1 en la formación de teobromina y teofilina, mientras que los estudios de proteínas recombinantes en microsomas no apoyan esto, sino que sugieren que contribuye a la formación de ácido 1,3,7-trimetilúrico (KOT). Los experimentos con microsomas han demostrado que CYP2C8, CYP2C9 y CYP3A4 también participan en el metabolismo primario de la cafeína (7).
El hábito tabáquico induce la actividad de CYP1A2, por tanto, el abandono del hábito de fumar en pacientes mientras están en tratamiento con un sustrato de CYP1A2, como la teofilina o la clozapina, aumenta sus concentraciones y puede provocar efectos adversos. Fumar también aumenta el aclaramiento de la cafeína debido a sus acciones sobre CYP1A2 y al dejar de fumar el aclaramiento inicial de cafeína disminuye significativamente (p<0,001) en un 36,1% (30,9% -42,2%), de 2,47 ml./min/kg de peso corporal (2,03-3,00 ml./min/kgde peso corporal) a un nuevo estado estable de 1,53 ml/min/kg de peso corporal (1,24-1,89 ml/min/kg de peso corporal) (8).
Farmacogenómica de la cafeína
Aspectos genómicos relacionados con el metabolismo hepático
Figura 4. Esquema de célula hepática con los genes
candidatos implicados en el metabolismo de la cafeína.

El alelo CYP1A2 * 1F es la variante más comúnmente estudiada con respecto a la cafeína. La variante que define este haplotipo es CYP1A2: (-163) C> A (rs762551). Un estudio de CYP1A2 * 1F, donde se excluyeron otros haplotipos que contienen (-163) C> A, mostró que (-163) AA se asoció con un aumento del metabolismo de la cafeína en fumadores suecos. En este estudio se observaron diferencias significativas en la actividad de la enzima CYP1A2, entre suecos y coreanos, que no pudieron ser explicadas por factores ambientales o los haplotipos CYP1A2 examinados, a pesar de las diferencias en las frecuencias alélicas. Ninguno de los haplotipos CYP1A2 investigados se mostró crítico para inducir variaciones en la actividad enzimática, con la excepción de CYP1A2 * 1F. Otros haplotipos que incluían (-163) C> A (* 1J, * 1K, * 21) no tenían un metabolismo significativamente alterado de la cafeína (9).
En un estudio de asociación del polimorfismo genético de CYP1A2 con el consumo de café, que comparaba población serbia y sueca, consumidores de elevadas cantidades de café, Sse observó una asociación significativa del consumo excesivo de café con una alta actividad de la enzima CYP1A2 solo en portadores de la variante genética -163 A/A. No se observaron diferencias significativas en la actividad de la enzima CYP1A2 entre los genotipos en los consumidores no intensos de café. Los resultados indican que el 22 y el 14% de la variabilidad fenotípica entre los grandes consumidores de café de Serbia y Suecia, respectivamente, podría explicarse por el polimorfismo -163C> A Concluyen los autores de este estudio que polimorfismo CYP1A2 -163C> A tiene un efecto creciente importante sobre la inducibilidad de CYP1A2 por el consumo excesivo de café y posiblemente sea un factor que contribuya a las variaciones interindividuales en la actividad de la enzima CYP1A2 (10).
Sin embargo, otro estudio del alelo CYP1A2 * 1K mostró una actividad CYP1A2 significativamente reducida en no fumadores en comparación con * 1A o * 1F, en población de origen etiope utilizando cafeína como sustrato de la sonda (11).
Aspectos genómicos relacionados con los receptores de adenosina
La variabilidad interindividual en el consumo de cafeína se atribuye, en parte, a diferencias individuales en respuesta a los efectos estimulantes de la cafeína. Ello, a su vez, dependería de un polimorfismo genético de los receptores de adenosina (12, 13)
Los estudios en gemelos han sugerido que los factores genéticos juegan un papel importante en la determinación de consumo habitual de cafeína y la respuesta a la misma. Sin embargo, los genes específicos involucrados no se han identificado.
En un estudio en gemelos idénticos realizado en Australia, las preferencias por el café parecieron estar relacionadas con factores genéticos (RR 0,42) y ambientales (RR 0,58). Se observó una relación inversa entre el consumo de té y café, mayor en las mujeres (-0,41) que en los hombres (-0,34). Esta asociación fue mediada enteramente por el ambiente en los varones, y tanto por el ambiente (68,3%) como por los genes (31,7%) en las hembras. El consumo de té y café tiene heredabilidades similares (0,46) en los hombres, pero el consumo de té se vio influenciado por factores ambientales comunes, mientras que el consumo de café no (14).
Los estudios de farmacogenómica, que examinan los efectos de las variantes en el receptor de adenosina ADORA2A y el comportamiento y las respuestas relacionados con la cafeína han tenido resultados variados.
En un estudio (Sobre 2735 individuos participantes en un estudio de interacciones gen-dieta y riesgo de infarto de miocardio) dirigido a examinar si la variabilidad genética de CYP1A2 (-163 A > C) o del receptor de adenosina A2A (ADORA2A 1083 C > T) se asocia con el consumo habitual de cafeína, se observó que el genotipo ADORA2A, pero no el CYP1A2, se asoció a diferentes cantidades de ingesta de cafeína.
La probabilidad de tener el genotipo ADORA2A 1083 TT disminuye a medida que aumenta el consumo habitual de cafeína, en comparación con otros genotipos del mismo receptor (CC + CT). Esta observación proporciona una base biológica para el comportamiento de consumo de cafeína y sugiere que las personas con este genotipo pueden ser menos vulnerables a la dependencia de la cafeína (15). Los genotipos ADORA2A (rs5751876 TT, rs2298383 CC y rs4822492 CC) se asociaron con aumento de la ansiedad en respuesta a la cafeína en una población sana sin un consumo regular excesivo (16)..
En personas que consumen habitualmente poca cafeína se ha señalado la asociación entre la variante rs5751876 del receptor ADORA2A y el nivel de ansiedad inducida por cafeína. Un estudio ha investigado si este polimorfismo de un solo nucleótido (SNP) pudiera relacionarse con la ingesta habitual de cafeína y si dicha ingesta habitual podría atenuar el efecto ansiogénico de la cafeína. Para ello se valoraron dos grupos de participantes, con diferentes niveles de consumo de cafeína, en forma de grupos paralelos, aleatorizados, doble ciego, que calificaron la ansiedad, el estado de alerta y el dolor de cabeza antes y después de 100 mg de cafeína y nuevamente después de otros 150 mg de cafeína, administrados 90 minutos después, o después del placebo en ambas ocasiones.
Los resultados mostraron una mayor susceptibilidad a ansiedad inducida por la cafeína, pero no se relacionó con un menor consumo (incluso este era mayor), el grupo de genotipo rs5751876 TT y una menor respuesta de ansiedad en relación con consumo alto/ medio versus consumo nulo/bajo, independientemente del genotipo. En este estudio, aparte del SNP rs3761422 de ADORA2A, casi completamente vinculado, no sebservó que ningún otro de los ocho SNP de ADORA2A y siete de ADORA1 estudiados estuviera claramente asociado con los efectos de la cafeína sobre ansiedad, estado de alerta o cefalea. La administración de placebo en los participantes con consumo alto o mediano disminuyó el estado de alerta y aumentó el dolor de cabeza. La cafeína no aumentó el estado de alerta en los participantes de con ninguno o bajo consumo.
Con el consumo frecuente, parece desarrollarse una tolerancia al efecto ansiogénico de la cafeína, incluso en individuos genéticamente susceptibles, pero no se obtiene ningún beneficio neto para el estado de alerta, ya que la abstinencia de cafeína reduce el estado de alerta y el consumo simplemente lo devuelve a la línea de base (17).
El genotipo CC de ADORA2A no se asocia con una respuesta vasodilatadora cuando se expone a adenosina o a cafeína (18). La distribución de distintos genotipos CC del gen del receptor de adenosina ADORA2A difiere entre adultos sensibles e insensibles a la cafeína. El genotipo CC de ADORA2A determina en qué medida los cambios inducidos por la cafeína en la actividad eléctrica del cerebro durante el sueño se asemejan a las alteraciones observadas en pacientes con insomnio. Estos datos demuestran un papel de los receptores de adenosina A2A para el sueño en humanos y sugieren que una variación común en ADORA2A contribuye a las respuestas subjetivas y objetivas a la cafeína durante el sueño (19).
Efectos farmacológicos de la cafeína
La cafeína atraviesa la barrera hematoencefálica y en el cerebro actúa como antagonista no selectivo del receptor de adenosina. La molécula de cafeína es estructuralmente similar a la adenosina, por lo que se une a los receptores de adenosina en la superficie de las células sin activarlos, actuando como un inhibidor competitivo. Las propiedades estimulantes de la cafeína dependen de su capacidad para reducir la transmisión de adenosina en el cerebro.
 |
Figura 5: Estructuras de las moléculas de cafeína, Adenosin monofosfato cíclico (AMPc) y Adenosina |
Tanto la cafeína como su principal metabolito, la paraxantina (1,7-dimetilxantina o 17X), inhiben de forma competitiva y no selectiva los receptores de adenosina in vitro. En un estudio de diseño cruzado, dirigido a valorar la contribución de la paraxantina a la actividad farmacológica de la cafeína (2 o 4 mg/kg decafeína oral versus placebo o paraxantina oral, a igual dosis que la cafeína, versus placebo, cada uno después de 3 días. de la abstinencia de metilxantina) se observó que, tanto la cafeína como la paraxantina aumentaron significativamente la presión arterial diastólica, los niveles de epinefrina plasmática y los ácidos grasos libres. La cafeína y la paraxantina produjeron una magnitud de respuesta similar a 4 mg/kg; sin embargo, la cafeína pareció producir mayores respuestas que la paraxantina a 2 mg/kg. Cafeína y la paraxantina tienen acciones simpaticomiméticas similares. La actividad de la paraxantina debe tenerse en cuenta para comprender la farmacología clínica de la cafeína, en particular con el consumo crónico y repetitivo de cafeína (20).
La cafeína antagoniza los receptores de adenosina (A1, A2A, A2B y A3), de forma no selectiva, que se expresan en los ganglios basales. Estos receptores aparecen en altas concentraciones en el núcleo accumbens y gánglios basales en el cuerpo estriado. Estas áreas forman parte de unas estructuras involucradas en varios aspectos del control motor y modulan la activación conductual y la toma de decisiones basadas en el esfuerzo.
Una evidencia creciente indica que el efecto estimulante psicomotor de la cafeína se genera al afectar a un grupo particular de neuronas de proyección ubicadas en el cuerpo estriado, el área principal de recepción de los ganglios basales. Estas células expresan altos niveles de receptores de ADORA2A, que están involucrados en varios procesos intracelulares, incluida la expresión de genes tempranos inmediatos y la regulación de la fosfoproteína DARPP-32 de 32 kDa regulada por dopamina y AMP cíclico (21).
La adenosina modula una variedad de funciones fisiológicas a través de la interacción con los receptores de adenosina A1 y A2, donde los agonistas median la inhibición y estimulación, respectivamente, de la adenilato ciclasa. En el sistema cardiovascular, los receptores A2 median la vasodilatación y la reducción de la presión arterial, mientras que los receptores A1 median la depresión cardíaca.
Los análogos de adenosina, en particular los compuestos sustituidos con N6, son más potentes en los receptores A1 que en los receptores A2. La subregión del receptor de adenosina que interactúa con el sustituyente N6 es diferente para los receptores A1 y A2, particularmente con respecto a las interacciones de fenilo, tolerancia en masa y estereoselectividad.
Las xantinas son antagonistas clásicos de los receptores de adenosina y muchas de sus acciones farmacológicas pueden deberse a ello. La cafeína y la teofilina son antagonistas no selectivas para los receptores A2. El reemplazo de los grupos metilo de la teofilina con n-propilo o grupos alquilo más grandes produce xantinas con selectividad para los receptores A1, particularmente cuando se combinan con un resto 8-fenilo.
Ciertos análogos de la cafeína en los que el grupo metilo en la posición 1 o 7 se reemplaza con un grupo propargilo o propilo muestran selectividad por los receptores A2 (22).
Da Chang de la Universidad Normal de Hangzhou en China y otros investigadores escanearon los cerebros de 60 participantes, 30 hombres y mujeres, al inicio del estudio y después de ingerir una pastilla de cafeína de 200 mg (aproximadamente dos tazas de café). El escaneo se realizó en reposo. Para ambas exploraciones, el equipo de Chang analizó los cambios en los niveles de actividad neuronal de un momento a otro y buscó correlaciones en la actividad a través y dentro de las regiones del cerebro para calcular la entropía cerebral. También midieron los cambios en el flujo sanguíneo cerebral.
La cafeína reduce el flujo sanguíneo cerebral, con un aumento de la entropía cerebral. Sin embargo, el aumento de entropía se correlacionó con la reducción del flujo sanguíneo solo en algunas regiones del cerebro. Los escáneres mostraron que la cafeína aumentaba la entropía cerebral especialmente en la corteza prefrontal lateral, la DMN (red de modo predeterminado, que involucra el soñar despierto y la autorreflexión), la corteza visual y la red motora. Los investigadores relacionaron los efectos beneficiosos conocidos de la cafeína en la función de atención, vigilancia y acción/movimiento. Hubo poca correlación local entre el aumento de la entropía y el flujo sanguíneo cerebral (que fue reducido por la cafeína), lo que sugiere que los efectos de la cafeína se debieron a influencias en la función neuronal, más que a cambios vasculares (23).
Otros efectos neurológicos de la cafeína han despertado un gran interés y permanecen en investigación. Como el efecto sobre la vascularización, que puede ser específico de una región y resultado de la distribución espacial de los receptores de adenosina (24). También se baraja la hipotesis de que la cafeína podría estimular la producción de cetonas como combustible alternativo para el cerebro durante la disminución de las concentraciones de glucosa (por ejemplo en la enfermedad de Alzheimer) (25). Además la cafeína puede afectar la calidad del sueño y aumentar el rendimiento en caso de privación del mismo. El EEG ha mostrado que reduce el sueño de ondas lentas, con un aumento de la vigilia y los despertares de la etapa 1 (26).
Propiedades antioxidantes de la cafeína
Para evaluar si la cafeína per se podría explicar las propiedades antioxidantes del café a corto plazo, se estudiado la capacidad de la ingestión de cafeína pura (5 mg/kg de peso corporal/día en dos dosis diarias durante siete días consecutivos) para mejorar los niveles plasmáticos de seis índices bioquímicos en voluntarios varones sanos (n = 15). Estos índices fueron la capacidad antioxidante total (TAC), glutatión (GSH), glutatión oxidado (GSSG), proporción de GSH a GSSG, hidroperóxidos de lípidos (LOOH) y malondialdehído (MDA). Encontrandose que todos los índices cambiaron significativamente (P <.05 o <.01) de manera favorable, oscilando entre -41% para GSSG y -70% para niveles de LHP, y + 106% para niveles de GSH a + 249% para Relación GSG / GSSG. Los cambios de cualquier índice dado fueron uniformes en todos los sujetos, sin valores atípicos. Concluyendo los autores ensayo que la cafeína tiene propiedades antioxidantes consistentes e inequívocas, sin valores atípicos (27).
Parece probado por la evidencia acumulada que la cafeína tiene propiedades antioxidantes, antiinflamatorias, anticolinesterasas y anti-TLR-4 (*).
(*) La activación de los toll like receptors (TLR) (CD284) juega un papel importante en el inicio de la respuesta inmune innata, incluso en pacientes críticos no sépticos. El eritoran, análogo del lípido A, actúa como antagonista de TLR4 y fue desarrollado (2009) como fármaco contra la sepsis grave. Sin embargo, los resultados contra la sepsis fueron algo decepcionantes y se utilizó con mejores resultados para tratar ciertos casos de influenza grave. Aunque no es un antiviral per se, podría usarse contra la reacción inmune masiva llamada tormenta de citocinas, que a menudo ocurre más tarde en la infección y es una de las principales causas de mortalidad por influenza grave.
Una mutación de pérdida de función espontánea en el gen TLR4 inhibe fuertemente la activación microglial y monocítica por el péptido amiloide de Alzheimer agregado, lo que resulta en una liberación significativamente menor de los productos inflamatorios IL-6, TNFalfa y óxido nítrico. El tratamiento de células neuronales primarias murinas con sobrenadante de microglia estimulada por péptido amiloide demuestra que Tlr4 contribuye a la neurotoxicidad microglial inducida por péptido amiloide (28).
En un estudio de 2019 se analizó el potencial antioxidante y antiinflamatorio de la cafeína en un modelo de ratón con neurodegeneración y deterioro sináptico inducidos por inyecciones de LPS (*). También notamos una mayor expresión del receptor 4 tipo toll (TLR4), el factor fosfo-nuclear kappa B (p-NF-kB) y la quinasa n-terminal fosfo-c-Jun (p-JNK) en los cerebros de ratones tratados con LPS, que se redujo significativamente en el grupo tratado conjuntamente con LPS + cafeína.
(*) El Lípido A es un componente lipídico de una endotoxina responsable de la toxicidad de las bacterias gramnegativas. Es la más interna de las tres regiones del lipopolisacárido (LPS) o endotoxina.
Los autores del estudio al que nos referimos concluyen que la cafeína previene el estrés oxidativo inducido por LPS en el cerebro del ratón. Además la cafeína suprime los mediadores y marcadores inflamatorios activados en células gliales, inducidos en el cerebro de ratón por LPS.También reduce la liberación de citoquinas en cerebro murino, así como la apostosis mitocondrial y la disfunción sináptica inducidas por LPS (29).
Los estudios en ratas han mostrado que estas propiedades juegan un papel esencial en la respuesta inflamatoria y el comportamiento ansioso inducidos por la privación del sueño.
Relación de los efectos de la cafeína con diversas patologías
Enfermedad de Alzheimer (EA)
En la fisiopatología de la EA, hay fenómenos como la agregación extracelular de péptidos β-amiloides (βA) y la proteína tau hiperfosforilada (ovillos neurofibrilares) pueden ser las causas más importantes de degeneración neural y del deficit cognitivo en la enfermedad (30). Además, la acumulación de datos ha implicado que la señalización de calcio desregulada puede tener una contribución importante a la muerte de las células neurales en la EA (31).
 |
Figura
6:
Vías amiloidogénicas y no amiloidogénicas del procesamiento de la
proteína precursora de β amiloide (βAPP). Los péptidos Aβ se
derivan del procesamiento de la βAPP a través de la vía
amiloidogénica, implicando el complejo β-secretasa (BACE1) y
γ-secretasa (compuesto de PS1 o PS2, Nicastrina, faringe anterior
defectuosa-1 (APH-1) y presenilina potenciador-2 (PEN-2)). La
producción de Aβ se elimina cuando el βAPP se procesa a través de
la vía no amiloidogénica que implica a los complejos de α-secretasa
y γ-secretasa. |
Varios sistemas neuronales, incluyendo N-Metl-D-Aspartato (NMDA), adenosina y rianodina (*), colaboran en el mantenimiento de la concentración intracelular de calcio en un estrecho rango (33, 34, 35).
(*) Los receptores de rianodina (RyR ) (un diterpenoide venenoso que se encuentra en la planta Ryania speciosa del grupo de las Salicaceae, originalmente usada como insecticida) forman una clase de canales de calcio intracelulares en varias formas de tejido animal excitable como músculos y neuronas. Hay tres isoformas principales del receptor de rianodina, que se encuentran en diferentes tejidos y participan en diferentes vías de señalización que involucran la liberación de calcio de los orgánulos intracelulares (RYR1, RYR2 y RYR3). Estos receptores son similares al receptor de trifosfato de inositol (IP3 ), y estimulados para transportar Ca2+ al citosol, reconociendo Ca2+ en el lado citosólico, estableciendo un mecanismo de retroalimentación positiva (36).
Los receptores RyR3 se expresan ampliamente en los tejidos, pero especialmente en el cerebro (37). Se ha demostrado que los niveles de receptores tipo RyR3 aumentan considerablemente en las células PC12 que sobreexpresan la presenilina 1 humana mutante, y en el tejido cerebral de los ratones knockin que expresan la presenilina 1 mutante a niveles normales. Ello parece indicar un papel en la patogenia de enfermedades neurodegenerativas, como la enfermedad de Alzheimer (38).
Por otra parte, los receptores de NMDA y adenosina parecen tener un papel en la fisiopatología y en el tratamiento de la EA. Lo que les convierte en posibles dianas terapéuticas (39).
Diferentes estudios han reportado alteraciones de la expresión y la función de los Receptores de RyR en cerebros humanos afectados por EA, en células que expresan mutaciones familiares ligadas a EA, en la proteína precursora de amiloide β (βAPP) y presenilinas (el núcleo catalítico en complejos de γ-secretasa que escinden la βAPP, generando así péptidos βA), así como en el cerebro de varios modelos de ratones AD transgénicos. Los datos sugieren que la expresión de RyR y la alteración funcional están asociadas a la patogénesis de la EA mediante el control de procesamiento de βAPP y producción de péptido βA; muerte neuronal; función sináptica y memoria y habilidades de aprendizaje (40).
 |
| Figura 7: Señalización neuronal Ca 2+ . Los aumentos de [Ca 2+ ] i citosólico son el resultado de un influjo a través de canales de Ca 2+ dependientes de voltaje (VGCC), receptores de glutamato ionotrópicos (receptores de ácido N-metil-D-aspártico, NMDAR; y alfa-amino -3-hidroxil-5-metil-4-isoxazol-propionato, AMPAR), y la liberación del RE a través de los receptores de inositol 1,4,5-trifosfato (IP 3 R) y rianodina (RyR). La compartimentación intraneuronal de Ca 2+ también se mantiene mediante la actividad de las proteínas tampón que se unen al Ca 2+ (p. ej., Calbindina y parvalbúmina), y se regula dentro de microdominios de señalización que involucran a l bomba ATP-Ca 2+ dependiente SERCA (retículo sarco-endoplásmico Ca 2+ ATPasa) acumulando Ca 2+ desde el citosol hacia ER, y el intercambiador sodio-Ca 2+ (Na+/Ca 2+ ), que actúan junto con PMCA (Plasma Membrane Ca 2+ ATPasa) para restaurar [Ca 2+ ] i de nuevo a los niveles de reposo mediante la extrusión de Ca 2+ del citosol al espacio extracelular. Aunque gran parte de la entrada de Ca 2+ en la neurona está mediada predominantemente por los canales de la membrana plasmática, la liberación de Ca 2+ mediada por IP 3 R y / o RyR se puede reclutar posteriormente a través del fenómeno de Ca inducido por Ca 2+Liberación 2+ (CICR), un proceso regenerativo en el que Ca 2+ mejora su propia liberación de IP 3 R y RyR. (40). |
Muchos casos de enfermedad de Alzheimer hereditaria (EA) de inicio temprano son causados por mutaciones en el gen de la presenilina-1 (PS1). Las mutaciones de PS1 pueden perturbar la homeostasis del Ca2+ celular y, por lo tanto, hacer que las neuronas sean vulnerables a la excitotoxicidad y la apoptosis. En ensayos en ratones knock-in mutantes para PS1, las células PC12 mutantes y neuronas primarias del hipocampo exhiben niveles muy aumentados de receptores de rianodina (RyR) y una mayor liberación de Ca2+ después de la estimulación con cafeína. Los análisis de inmunotinción y co-inmunoprecipitación de marcaje doble indican que PS1 y RyR están colocalizados e interactúan físicamente (Figura 5). El tratamiento con cafeína sensibiliza a las neuronas que expresan PS1 mutante a la apoptosis inducida por el péptido βA, vinculado a la patogenia de la EA.
 |
| Figura 8: Las proteínas PS1 y RyR se colocalizan e interactúan. A , micrografía de escaneo láser confocal que muestra una imagen combinada (anaglifo) de células PC12 marcadas doblemente con anticuerpos PS1 y RyR. Rojo , inmunorreactividad de PS1; verde , inmunorreactividad RyR; amarillo , sitios de colocalización. Chan S L et al. (38). |
Los niveles de RyR3 aumentan considerablemente en las células PC12 que sobreexpresan la PS1 humana mutante y en el tejido cerebral de los ratones knock-in que expresan la PS1 mutante a niveles normales. Los estudios de imágenes de calcio mostraron que las células PC12 y las neuronas corticales que expresan PS1 mutante exhiben respuestas de calcio aumentadas a la cafeína en comparación con las células que expresan PS1 de tipo salvaje (38).
Los mecanismos subyacentes a la disfunción del receptor de rianodina (RyR), asociada con la EA aún no se comprenden bien. Aunque se ha sugerido que los canales RyR2 neuronales experimentan remodelación postraduccional (fosforilación, oxidación y nitrosilación de PKA) en el cerebro de pacientes con EA. Este fenómeno postraduccional se ha reproducido experimentalmente en dos modelos murinos de EA. Los ratones genéticamente modificados en sus receptores RyR2, exhibieron fugas congénitas de CA2+ en el sistema reticulo-endoplasmático (SRE), presentando defectos prematuros y severos en la plasticidad sináptica, así como en el comportamiento y la función cognitiva. Estos datos proporcionan un mecanismo subyacente a los canales RyR2 con fugas, que podrían considerarse como posibles dianas terapéuticas para la EA (41,42).
La evidencia acumulada confirma que la cafeína produce un efecto neuroprotector contra la neurotoxicidad de la proteína βA sobre líneas celulares neuronales. Tanto el bloqueo de los receptores de adenosina y de N-Metl-D-Aspartato (NMDA), como la activación de receptores de rianodina (RyR), pueden contribuir a los efectos neuroprotectores de la cafeína.
Los estudios epidemiológicos también confirman la neuroprotección mediada por cafeína y revelan una incidencia de EA inversamente proporcional al consumo de cafeína (43,44). Sin embargo, algunos estudios han comunicado que la administracion aguda de cafeína puede tener un efecto deletéreo sobre sobre el estado neurológico (45). Las inconsistencias en los resultados de estudios sobre los efectos neuroprotectores de la cafeína pueden ser atribuibles a diferencias metodológicas en los ensayos.
Los resultados de algunos estudios han mostrado que, diferentes concentraciones de cafeína protegen significativamente las células neuronales contra la neurotoxicidad βA . En dicho estudio se evaluó también la interacción de adenosina, NMDA y dantroleno (antagonista del receptor de rianodina), con los efectos neuroprotectores de la cafeína, con dos concentraciones de la misma (0,6 y 1 mM). La cafeína en todas las concentraciones utilizadas (0,6, 0,8, 0,9, 1 y 3 mM) protegió significativamente a las células neuronales contra la neurotoxicidad Aβ. La adenosina a las concentraciones de 20, 40, 80 y 100 μM disminuyó los efectos neuroprotectores de la cafeína (0,6 y 1 mM) contra la neurotoxicidad Aβ. El NMDA a las concentraciones de 20, 50, 70 y 90 μM bloqueó los efectos neuroprotectores de la cafeína (0,6 y 1 mM). El dantroleno a las concentraciones de 2, 4, 6, 8 y 10 μM disminuyó los efectos neuroprotectores de la cafeína (0,6 mM) y a las concentraciones de 2 y 10 μM impiden la neuroprotección de la cafeína (1 mM) contra la neurotoxicidad Aβ (46).
Los RyR se activan mediante concentraciones milimolares de cafeína. Concentraciones de cafeína superiores a 5 mmol/L provocan un aumento pronunciado en la sensibilidad de los RyR al Ca2+, haciendo que las concentraciones basales de Ca2+ se vuelvan activantes. A concentraciones bajas de cafeína milimolar, el receptor se abre de forma cuántica, pero tiene un comportamiento complicado en términos de uso repetido de cafeína o dependencia de las concentraciones de calcio citosólico o luminal.
Los estudios en ratas mostraron que estas propiedades juegan un papel esencial en la respuesta inflamatoria y el comportamiento ansioso inducidos por la privación del sueño.
Enfermedad de Parkinson
Se ha confirmado repetidamente una relación entre el aumento de la ingesta de cafeína y un menor riesgo de enfermedad de Parkinson (EP).
En modelos animales de EP, inducida por 1‐metil‐4‐fenil 1,2,3,6‐tetrahidropiridina, la cafeína protege las neuronas al bloquear el receptor de adenosina ADORA2A. La cafeína es metabolizada principalmente por el citocromo P450 1A2 (CYP1A2). Se han estudiado los polimorfismos de los genes ADORA2A y CYP1A2 en su asociación con el riesgo de EP y su influencia sobre la asociación cafeína-EP.
Basados en el Parkinson’s Epidemiology and Genetic Associations Studies in the United States (PEGASUS), sobre epidemiología y asociaciones genéticas de EP en EE.UU., que incluyeron cinco estudios de caso-control basados en la población, se genotiparon cuatro polimorfismos de ADORA2A y tres de CYP1A2 en 1325 casos de EP y 1735 controles emparejados por edad y sexo. La información sobre el consumo de cafeína se obtuvo mediante entrevistas estructuradas en persona o telefónicas.
Dos polimorfismos de ADORA2A se asociaron inversamente con el riesgo de EP: rs71651683, una variante de 5 ' (OR alélico ajustado = 0,51, IC del 95% 0,33-0,80, p=0,015 ajustado por permutación) y rs5996696, una variante de la región promotora (La OR, ajustado para genotipos AC y CC, comparados con el genotipo AA salvaje, fueron 0,76 (IC del 95%: 0,57–1,02) y 0,37 (IC del 95%: 0,13–1,01), respectivamente (p ajustada por permutación para la tendencia = 0,04). Los polimorfismos de CYP1A2 no se asociaron con riesgo de EP; sin embargo, la asociación café-PD fue más fuerte entre sujetos homocigotos para el alelo variante rs762551 (interacción p=0.05) o rs2470890 (interacción p=0.04).
En este gran estudio, dos polimorfismos de ADORA2A se asociaron inversamente con el riesgo de EP, pero hubo evidencia débil de interacción con el consumo de café. Por el contrario, la asociación café-PD fue más fuerte entre los metabolizadores lentos de la cafeína que eran portadores homocigotos de los polimorfismos CYP1A2 (47).
Se ha demostrado que los niveles de cafeína y nueve de sus metabolitos son significativamente más bajos en las personas con EP, en comparación con las que no la padecen. Este hallazgo no dependió de la ingesta total de cafeína. Aunque ambos grupos tomaron un promedio de aproximadamente 2 tazas de café por día, aquellos con EP tenían un nivel total de cafeína de 24 pmol/10 µL, en comparación con 79 pmol/10 µL para los voluntarios sanos (48)
Además, hubo una tasa de sensibilidad del 77% y una tasa de especificidad del 74% para la cafeína en muestras de suero, como biomarcador de diagnóstico para la EP. Se ha sugerido que esos hallazgos pueden representar biomarcadores viables para la identificación de la enfermedad de Parkinson temprana.
Los investigadores inscribieron a 108 pacientes con EP de leve a moderada pero sin demencia (54% hombres; edad media, 67,1 años) en un hospital de Japón entre diciembre de 2013 y febrero de 2014. El grupo de comparación de edad comprendía 31 adultos sanos (43% hombres ; edad media, 63,3 años). Se utilizó cromatografía líquida y espectrometría de masas para medir los niveles de cafeína y los 11 metabolitos de las muestras de suero sanguíneo de todos los participantes.
Los resultados mostraron que los niveles generales de cafeína fueron significativamente más bajos en los pacientes con EP que en aquellos sin EP (p<0,0001). Nueve de los 11 metabolitos de la cafeína también fueron significativamente más bajos para el grupo de EP, incluidos teofilina (p<0,0001), teobromina (p= 0,0004) y paraxantina (p<0,0001) (48).
Algunos investigadores han sugerido que probablemente, las personas con riesgo de EP podrían tener malabsorción de cafeína en intestino delgado. Ello podría plantear el uso de la vía transdérmica, en el caso de valorar una terapia con cafeína. Considerándose que la ingesta alta de cafeína pudiera detener la progresión de la enfermedad (49).
Un estudio reciente se centró específicamente en los efectos de la cafeína en individuos con una mutación en el gen de la quinasa repetida rica en leucina 2 (LRRK2). El estudio encontró que ese grupo en particular puede beneficiarse de las terapias relacionadas con la cafeína. Se ha demostrado que quienes padecen la enfermedad de Parkinson tienen niveles más bajos de cafeína que quienes no la padecen; los individuos con mutaciones LRRK2 tenían niveles incluso más bajos que los de otros pacientes con enfermedad de Parkinson (50).
Están menos claros los efectos de la cafeína sobre los síntomas motores en pacientes con enfermedad de Parkinson son menos claros. Aunque algunos hallazgos han sugerido que la ingesta de cafeína mejoró la marcha y otras preocupaciones, la cafeína administrada en una dosis equivalente a tres tazas de café al día no logró mejorar los síntomas motores.
Aunque la cafeína puede ayudar en el riesgo y la progresión de la enfermedad de Parkinson, la levodopa sigue siendo recomendada como tratamiento de primera línea para los síntomas motores (51).
Cefaleas
La cafeína ha demostrado su utilidad, sola o asociada a otros fármacos, en el tratamiento del dolor de cabeza. La FDA reconoció la combinación de ácido acetilsalicílico (250 mg), acetaminofén (250 mg) y cafeína (65 mg) como segura y eficaz para tratar los dolores de cabeza agudos, especialmente la migraña, y también fue considerada eficaz por la American Headache Society (Nivel A ). Esta combinación se tolera bien en la cefalea tensional episódica y se considera superior al acetaminofén solo. Todos los componentes de esta combinación se consideran seguros durante la lactancia y se pueden tomar por vía oral para los ataques de migraña aguda (52).
Además de las migrañas y los dolores de cabeza de tipo tensional, la cafeína se considera eficaz para otros tipos de dolores de cabeza, como el dolor de cabeza posterior a la punción dural y los dolores de cabeza hipnícos (a los que nos referimos mas adelante), que pueden presentarse con características autonómicas craneales.
Los estudios clínicos mostraron que el citrato de cafeína (intravenoso [IV]) podría usarse solo en los ataques de migraña aguda, pero podría ser inferior al sulfato de magnesio (IV) en las migrañas de moderadas a severas. La administración temprana de compuestos de ergotamina/cafeína también puede ser eficaz. Un estudio clínico mostró que suspender la cafeína antes de un ataque de migraña puede aumentar la eficacia del tratamiento agudo.
La cefalea hípnica (CH) es un tipo de cefalea poco frecuente, benigna, descrita por primera vez por Raskin en 1988 (54), que se caracteriza por episodios nocturnos recurrentes que despiertan periódicamente a los pacientes dormidos y se observa predominantemente en mayores de 50 años. Sin embargo, las personas más jóvenes e incluso niños pueden experimentar dolor de cabeza de carácter hípnico. Los ataques suelen durar entre 15 y 180 minutos, pero algunos pacientes informan ataques de dolor de cabeza de hasta 10 horas. Casi todos los pacientes informan actividad motora durante los ataques de dolor de cabeza. Se ha demostrado que la cafeína produce la mejor respuesta terapéutica en el tratamiento agudo (53)
En 2019 se publicó una revisión que tuvo como objetivo describir las características clínicas de los casos de CH publicados en la literatura desde 1988 a 2018. El estudio se basó en la búsqueda de literatura en las principales bases de datos médicas (LiLacs, SciELO, Bireme, Medline, Embase, Current Contents, Scopus, EBSCO y PubMed) y el análisis de los casos clínicos de CH publicados. El estudio incluyo a 343 adultos (69,0% mujeres y 31,0% hombres) y 5 niños (3 niñas y 2 niños) diagnosticados de CH. La edad promedio para adultos y niños fue, respectivamente, 58.0 ± 13.1 años (rango de 15 a 85 años) y 9 años (rango de 7 a 11 años). El diagnóstico se realizó a 7,6 ± 14,2 años (rango de 0,1 a 39 años) después del inicio de la cefalea. El dolor se presentó durante el sueño nocturno (94,8%), con una duración media de 90 min, de localización bilateral (55,5%), de carácter sordo (74,4%) e intensidad moderada (61,5%). En el 94,5% de los pacientes, el dolor de cabeza se produjo durante 10 o más días al mes (media de 21 días). Las manifestaciones autonómicas ocurrieron en el 7,6% de los pacientes, predominantemente lagrimeo (61,1%) y rinorrea (16,7%) (54)
La cafeína presentó la mejor respuesta terapéutica en el tratamiento agudo. En profilaxis, el litio, la cafeína y la indometacina fueron fármacos eficaces en el 77,8% de los pacientes. En el 56,7% de los pacientes hubo remisión con tratamiento y en el 72,7% sin recidiva. La CH es una enfermedad rara que generalmente ocurre por primera vez en mujeres mayores, pero puede comenzar en la niñez (54).
Los medicamentos para el dolor de cabeza con cafeína, ya sea solos o en combinación, son ampliamente utilizados (Tabla 2). Los médicos deben estar familiarizados con su uso, así como con la química, la farmacología, las fuentes dietéticas y médicas, los beneficios clínicos y los posibles problemas de seguridad de la cafeína.
En una revisión de 2017, en comparación con la medicación analgésica sola, las combinaciones de cafeína con medicamentos analgésicos, incluidos acetaminofén, ácido acetilsalicílico e ibuprofeno, mostraron una eficacia significativamente mejorada en el tratamiento de pacientes con CTT o migraña, con buena tolerancia en la gran mayoría de los pacientes. Los eventos adversos más comunes fueron nerviosismo (6,5%), náuseas (4,3%), dolor / malestar abdominal (4,1%) y mareos (3,2%). Esta revisión proporciona evidencia de la función de la cafeína como coadyuvante en el tratamiento agudo de la cefalea primaria con otros fármacos de uso común. Dosis de cafeína de 130 mg mejoran la eficacia de los analgésicos en la CTT y las dosis de ≥100 mg mejoran los beneficios en la migraña. Los autores señalan que se necesitan estudios adicionales para evaluar la relación entre la dosis de cafeína y los beneficios clínicos en pacientes con CTT y migraña (55).
Hemorragia subaracnoidea y hemorragia intracerebral
La cafeína es el neuroestimulante central de uso mas extendido en el mundo. Se encuentra en muchos alimentos y bebidas (particularmente en el café, el té y algunos refrescos y estimulantes). También se encuentra en algunos fármacos, asociada otros principios activos, contribuyendo a realzar sus efectos farmacológicos. Es frecuente la automedicación y en muchos casos los consumidores ignoran el contenido de cafeína como ingrediente (tanto en fármacos como en bebidas estimulantes).
En cualquier caso, la principal fuente de cafeína para la población es el café. Por ello la mayoría de los estudios en la literatura médica están dedicados a la relación entre el consumo de café y el riesgo de enfermedad.
Existe una creciente evidencia de que el consumo de café no incrementa el riesgo de de ictus hemorrágico ni de otros eventos cardiovasculares mayores como la isquemia coronaria. Sin embargo, existen datos contradictorios en la literatura. En un estudio de 1990, que examinaba prospectivamente la relación entre el consumo de café y el riesgo de infarto de miocardio (IM) (que necesitó de intervención con by-pass o angioplastia coronaria) y de ictus, en una cohorte de 45.589 hombres de EE.UU. de entre 40 a 75 años, sin historia de enfermedad cardiovascular, se observó que el consumo total de café no se asoció con un aumento del riesgo. Sin embargo se observó un incremento marginalmente significativo del riesgo en pacientes consumidores de café descafeinado. Además no se observó en este estudio un incremento del riesgo en subgrupos de pacientes que tenían un aumento de ingesta de cafeína procedente de otras fuentes. Y, por otra parte, el ajuste realizado por indicadores de riesgo cardiovascular mayores e ingesta de grasas y colesterol en la dieta no supuso una alteración de las asociaciones observadas (56).
En otro sentido, en una cohorte de hombres, inscritos en el Honolulu Heart Program, y seguidos durante un periodo de 25 años para estudio de la incidencia ictus, de los cuales 499 eran hipertensos (con una presión arterial sistólica o diastólica mayor de 140 y 90 mmHg, respectivamente), no fumadores y con edades comprendidas entre 55 y 68 años, se observó una asociación positiva del consumo de café con el aumento de riesgo de ictus isquémico (57).
En un estudio de seguimiento prospectivo de 3.837 pacientes finlandeses seleccionados al azar con DM2, de entre 25 y 74 años, para identificar casos de cardiopatía coronaria (EC), ECVA e ictus, se valoró el consumo de café y otros parámetros al inicio del estudio. Las asociaciones entre el consumo de café al inicio del estudio y el riesgo de mortalidad total, ECV, CC e ictus se analizaron mediante el uso de modelos de riesgos proporcionales de Cox (58).
 |
Tabla
3:
HR (IC del 95%) de la mortalidad total, por ECVA, EC
e
ictus
por volumen de café como variable continua y por diferente volumen
de café (≥3 tazas frente a 0-2 tazas) entre sujetos con DM2 (58). |
Se observó un riesgo reducido de mortalidad total, ECVA y EC entre los pacientes que bebían 2 tazas o más de café al día. Esta asociación fue relativamente fuerte y no se vio afectada por los otros factores modificadores del efecto incluidos en los análisis. Después de ajustar por los otros factores de riesgo potenciales de ECVA, el resultado incluso se volvió algo más fuerte. Sin embargo, esta asociación de riesgo inversa con el consumo de café no pudo demostrarse en relación con el ictus (58).
En un estudio prospectivo realizado sobre una cohorte de hombres fumadores, también en población finlandesa, en este caso reclutada del Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study, se observó una asociación inversa significativa, del consumo de té y café, con el riesgo de ictus isquémico. La comparación entre participantes en el estudio con consumo bajo o no consumidores de estas bebidas y los que consumían 8 o mas tazas de café al día y 2 o mas tazas de té al día, dio como resultado un descenso del riesgo de ictus del 23 y 21 % respectivamente. Esta asociación fue independiente de otros factores de riesgo conocidos (59).
Otro estudio prospectivo, sobre la asociación de consumo de café y riesgo de ictus, se basó en una cohorte de 83.076 mujeres, incluidas en el Nurses` Health Study (NHS), de la que se excluyeron para su análisis participantes con antecedentes de ictus, EC, diabetes o cáncer, así como aquellas de las que no constaba información sobre el consumo de café. El seguimiento se prolongó durante 24 años (desde 1980 a 2004), con valoración del consumo de café a lo largo del tiempo y cada 2 a 4 años. Durante el seguimiento se documentaron 2280 ictus, de los cuales 426 fueron hemorrágicos, 1224 isquémicos y 630 indeterminados (60).
Utilizando un modelo de regresión de Cox, con ajuste por edad, consumo de tabaco, IMC, actividad física, ingesta de alcohol, menopausia, terapia hormonal, consumo de aspirina y factores dietéticos, los riesgos relativos (RR) de ictus, en relación con las diferencias categorías de consumo de café (<1 taza al mes; de 1 al mes a 4 semanales; de 5 a 7 semanales; 2 a 3 diarias; y ≥4 al día), fueron 1, 0.98 (95% CI, 0.84 a 1.15), 0.88 (95% CI, 0.77 a 1.02), 0.81 (95% CI, 0.70 a 0.95) y 0.80 (95% CI, 0.64 a 0.98) (P por tendencia=0.003). Tras un ajuste posterior en función de hipertensión arterial, hipercolesterolemia y DM2 se mantuvo la asociación inversa entre consumo de café y riesgo de ictus.
Por tanto, el consumo prolongado de café, a lo largo de los años, no se asocia con un aumento del riesgo de ictus en mujeres y los datos sugieren que el consumo de café puede generar una modesta reducción del riesgo de ictus. Tampoco se asoció con ictus el consumo de té o refrescos con cafeína. Y el consumo de café descafeinado también se asoció con una tendencia a menor riesgo de Ictus, después de ajuste para el consumo de café con cafeína (60).
La investigación sobre la relación entre los medicamentos que contienen cafeína (MCC) y el riesgo de ictus hemorrágico (IH) es escasa. Un estudio de 2013 evalúa la asociación entre los MCC y el riesgo de ictus hemorrágico.
Se trata de un estudio multicéntrico caso-control realizado en Corea del Sur, de 2002 a 2004. Un total de 940 pacientes con IH agudo, de 30 a 84 años, sin antecedentes previos de ictus, 940 comunitarios y 940 controles hospitalarios, pareados por edad y sexo en cada caso. Se registraron todos los fármacos tomados en los 14 días anteriores a la fecha (fecha índice) y la hora del inicio del accidente cerebrovascular (tiempo cero) para los sujetos de casos o el tiempo cero emparejado para los sujetos de control. La exposición a los MCC se definió por el uso de los msmos en la fecha índice antes del inicio o durante los 3 días anteriores. Los RR ajustados y sus intervalos de confianza (IC) del 95% se estimaron mediante regresión logística condicional.
Los resultados sugerían que el uso de CCM se asocia con un mayor riesgo de IH, tanto hemorragia subaracnoidea como hemorragia intracerebral. Curiosamente, esta asociación era mas significativa entre los pacientes que no tomaban café habitualmente (61)
Hepatopatía crónica
Una revisión sistemática encontró que los pacientes con enfermedad hepática crónica que consumen café tienen un menor riesgo de progresión a cirrosis, una menor tasa de mortalidad y una menor tasa de desarrollo de carcinoma hepatocelular.
Para examinar la asociación del consumo de café con la enfermedad hepática, se realizó una revisión sistemática de los estudios sobre los efectos del café en las pruebas de laboratorio asociadas al hígado, la hepatitis viral, la enfermedad del hígado graso no alcohólico (EHGNA), la cirrosis y el carcinoma hepatocelular (HCC). El consumo de café se asoció con mejores valores séricos de gamma glutamiltransferasa, aspartato aminotransferasa y alanina aminotransferasa de manera dependiente de la dosis en personas con riesgo de enfermedad hepática.
En pacientes con enfermedad hepática crónica que consumen café, se observó un menor riesgo de progresión a cirrosis, una tasa de mortalidad más baja en pacientes con cirrosis y una tasa más baja de desarrollo de HCC. En pacientes con hepatitis C crónica, el café se asoció con mejores respuestas virológicas a la terapia antiviral. Además, el consumo de café se relacionó inversamente con la gravedad de la esteatohepatitis en pacientes con enfermedad del hígado graso no alcohólico. Por tanto, en pacientes con enfermedad hepática crónica, es beneficioso el consumo diario de café (62).
Mas recientemente (2017) (63) un metanálisis de 16 estudios, que incluyeron más de 3,000 bebedores de café y 132,000 no consumidores, que tenía como objetivo examinar la asociación entre el café, incluyendo café con cafeína y descafeinado, con el carcinoma hepatocelular (CHC) y evaluar la influencia de la etiología del mismo y de la enfermedad hepática preexistente.

Además, los investigadores encontraron que el consumo de café también se asoció con una reducción del 27% en la probabilidad de desarrollar fibrosis hepática avanzada, en comparación con la de los no bebedores de café (63).
El estudio concluyó que los bebedores de café tenían un 39% menos de probabilidades de desarrollar cirrosis. Además hubo evidencia de un efecto dosis-respuesta: los consumidores de dos o más tazas diarias tuvieron una reducción del 47% en el riesgo de cirrosis, en comparación con los no bebedores, mientras que un consumo más modesto se asoció con una reducción del 34%. independientemente de la etapa de la enfermedad hepática del paciente o la presencia o ausencia de alto consumo de alcohol, tabaquismo, obesidad, diabetes tipo 2 o infección por hepatitis B o C.
Arritmia cardiaca
Hay muchos factores conocidos relacionados con con aumento del riesgo de fibrilación auricular (FA), entre los que se encuentran la hipertensión arterial (HTA), la diabetes mellitus, la insuficiencia cardíaca, la obesidad, hipertiroidismo o la cardiopatía isquémica. Sin embargo hay pocos datos sobre la influencia de los hábitos alimentarios sobre el riesgo de desarrollar FA.
El café es una infusión de consumo muy frecuente en occidente, que contiene cafeína, polifenoles y terpenos, sustancias que han demostrado influencia sobre la patología cardiovascular (64). Entre sus efectos fisiológicos conocidos están la liberación de adrenalina, noradrenalina y renina (65). Además aumenta la sensibilidad del miocardio a Ca++ (66,67), lo que puede influir sobre el ritmo cardiaco.
Algunos estudios realizados a cerca de la relación del consumo de café con el riesgo de FA han arrojado resultados contradictorios y poco significativos (68,69).
La revisión de los efectos cardiovasculares en pacientes aquejados de diferentes patologías cardiacas (hipertensión arterial, enfermedad arterial coronaria, alteraciones del ritmo e insuficiencia cardíaca) ha mostrado que la ingesta moderada de té o café no parece tener un efecto deletéreo sobre estas patologías e incluso puede ser beneficiosa en algunos casos, incluido la enfermedad coronaria, la insuficiencia cardíaca y las arritmias (70).
En 2019 se publicó un estudio prospectivo realizado en EEUU, en 18960 hombres, con una edad media de 66,1 años, participantes en el Physicians Health Study. Se evaluó el consumo de café mediante cuestionarios de frecuencia alimentaria autoinformados. La incidencia de FA se valoró mediante cuestionarios anuales, que fueron validados mediante revisión de historias clinicas en una submuestra de los pacientes (71).
Durante un seguimiento medio de 9 años se detectó aparición de 2098 nuevos casos de FA. Las relaciones de riesgo (95% IC) de FA fueron 1.0 (referencia), 0.85 (0.71‐1.02), 1.07 (0.88‐1.30), 0.93 (0.74‐1.17), 0.85 (0.74‐0.98), 0.86 (0.76‐0.97), y 0.96 (0.80‐1.14), para un consumo de café variable: consumo raro o ninguno, ≤1 taza/semana, de 2 to 4 tazas/semana, 5 to 6 tazas/semana, 1 taza/día, de 2 a 3 tazas/día and 4 o mas tazas/día, respectivamente. Resultados ajustados por edad, tabaquismo, consumo de alcohol y ejercicio físico. Los resultados de este estudio sugieren menor riesgo de FA entre aquellos hombres que consumieron entre 1 y 3 tazas de café al día (71).
El consumo diario moderado de café no tuvo un efecto adverso aparente para desencadenar arritmias cardíacas incidentes, e incluso se relacionó con una caída pequeña pero estadísticamente significativa de las arritmias en un análisis de datos recopilados prospectivamente de casi 300,000 residentes del Reino Unido. Dada la inconsistencia de los resultados de muchos de los estudios antes citados, este estudio de 2020 se planteó como objetivo investigar la asociación entre el consumo habitual de cafeína y el riesgo de arritmia en un gran estudio prospectivo de cohorte basado en la población (72).
Se valoró la información disponible, en relación al consumo de café con diferentes variables relevantes, a partir de los datos procedentes de UK Biobank, para 357.022 participantes con una edad media de 56,83 años y un 53,5 % de mujeres. Se realizó un análisis de regresión de riesgo (Cox multivariante), para probar la asociación entre consumo de café y el riesgo de arritmias.
Durante 5,25 ± 2,1 años, se diagnosticaron 8.159 arritmias incidentes (6.999 fibrilación o flutter auricular, 890 taquicardias supraventriculares, 459 taquicardias ventricularer, 385 complejo ventricular prematuro). En comparación con el no consumo, el consumo de café de 1-2, 3-4, 5 o más tazas al día se asoció con un riesgo significativamente menor de arritmia (HR 0,90, p <0,0001; HR 0,86, p <0,0001; HR 0,85 p = 0,0005 ; HR 0.88, P = 0.05, respectivamente, Figura XX). Cada taza de café diaria adicional se asoció con un 4% menos de incidencia de arritmia (HR 0,96, IC 0,941-0,98, p <0,0001). Se observó una reducción en la incidencia de arritmias para cada subtipo, excepto para los complejos ventriculares prematuros, que no alcanzaron significación estadística. La relación se mantuvo para las personas que informaron haber bebido hasta cinco o seis tazas de café al día (72).
 |
Figura 9: Incidencia de arritmia en relación al consuno habitual de café (72) |
Los autores concluyen que el consumo regular de café se asocia con un riesgo significativamente menor de arritmias (72).
Kim EJ, Nah G, Vittinghoff E, Delling FB, Marcus GM. Is drinking coffe associated with an increased risk of arrhythmia? Heart Rhythm Society (HRS) 2020: Póster D-PO01-032.
Estos resultados se suman a una base sustancial de evidencia existente que documenta la seguridad del consumo moderado y habitual de café cuando se trata de ritmos cardíacos
Enfermedad renal crónica (ERC)
Con el objetivo de aclarar la asociación de consumo de cafeína y mortalidad en pacientes con ERC y, dada la escasez de datos relevantes a respecto en la literatura, se realizó un gran estudio observacional en el que se analizaron los datos de 4863 adultos estadounidenses, no institucionalizados con ERC (definida por una tasa de filtración glomerular (eGFR) de 15 a 60 mL/min/1,73 m² y/o un cociente albumina/creatinina>30 mg/g) dentro de un estudio nacional, utilizando el National Health and Nutrition Examination Survey (NHANES) 1999–2010 (73).
En este grupo el consumo de cafeína se evaluó mediante encuestas dietéticas en las 24 h anteriores y se evaluó la mortalidad por todas las causas, cardiovascular y por cáncer, hasta el 31 de diciembre de 2011. También se realizó un análisis del consumo de cafeína según su origen (café, té y refrescos). Los cuartiles de consumo de cafeína fueron <28,2 mg/día (Q1), 28,2-103,0 (Q2), 103,01-213,5 (Q3) y >213,5 (Q4).
Se ha observado un asociación inversa entre la cafeína y mortalidad por todas las causas entre los participantes con ERC. No detectamos una asociación significativa entre el consumo de cafeína y la mortalidad cardiovascular o por cáncer. No hubo interacciones significativas entre los cuartiles de consumo de cafeína y las etapas de la ERC o las categorías de relación albúmina/creatinina en orina con respecto a la mortalidad por todas las causas. En consecuencia, el consumo de cafeína parece ser seguro en diferentes etapas de la enfermedad renal.
El equipo de investigación también encontró una relación inversa inesperada entre el consumo de cafeína a través de refrescos y la mortalidad por todas las causas en pacientes con ERC.
Teniendo en cuenta el contenido medio de cafeína por unidad de bebida con cafeína, incluso pequeñas cantidades de bebidas de consumo común pueden conferir un efecto protector con respecto a mortalidad por todas las causas en pacientes con ERC, concluyen los investigadores. El mecanismo que confiere el efecto protector del consumo de cafeína es incierto.
Es posible que el mismo mecanismo que justifica la relación inversa entre el consumo de café y la mortalidad reportada en la población general también exista con la cafeína en pacientes con ERC (73).
- https://www.ocu.org/alimentacion/cafe/calculadora/cafeina
- Adan A, Prat G, Fabbri M, Sànchez-Turet M (2008-10). Early effects of caffeinated and decaffeinated coffee on subjective state and gender differences..Progress in Neuro-Psychopharmacology and Biological Psychiatry 2008-10; 32(7):1698-1703.
- Liguori A, Hughes JR, Grass JA. Absorption and Subjective Effects of Caffeine from Coffee, Cola and Capsules. Pharmacol Biochem Behavior 1997; 58(3):721-726.
- Newton R, Broughton LJ, Lind MJ, Morrison PJ, Rogers HJ, Bradbrook, ID. Plasma and salivary pharmacokinetics of caffeine in man. European Journal of Clinical Pharmacology 1981; 21(1):45-52.
- Begas E, Kouvaras E, Tsakalof A, Papakosta S, Asprodini EK. In vivo evaluation of CYP1A2, CYP2A6, NAT-2 and xanthine oxidase activities in a Greek population sample by the RP-HPLC monitoring of caffeine metabolic ratios. Biomed. Chromatogr. 2007; 21:190–200.
- Kot M, Daniel WA. Caffeine as a marker substrate for testing cytochrome P450 activity in human and rat. Pharmacol Rep, 2008; 60(6):789-97.
- Kot M, Daniel WA. The relative contribution of human cytochrome P450 isoforms to the four caffeine oxidation pathways: an in vitro comparative study with cDNA-expressed P450s including CYP2C isoforms. Biochem Pharmacol, 2008; 76(4):543-51.
- Faber MS, Fuhr U. Time response of cytochrome P450 1A2 activity on cessation of heavy smoking. Clin Pharmacol Ther, 2004; 76(2):178-84.
- Ghotbi R, Christensen M, Roh HK, Ingelman-Sundberg M, Aklillu E, Bertilsson L. Comparisons of CYP1A2 genetic polymorphisms, enzyme activity and the genotype-phenotype relationship in Swedes and Koreans. Eur J Clin Pharmacol 2007; 63(6):537-46.
- Djordjevic N, Ghotbi R, Jankovic S, Aklillu E. Induction of CYP1A2 by heavy coffee consumption is associated with the CYP1A2 -163C>A polymorphism. Eur J Clin Pharmacol 2010; 66(7):697-703
- Aklillu E, Carrillo JA, Makonnen E, Hellman K, Pitarque M, Bertilsson L, Ingelman-Sundberg M. Mol Pharmacol 2003; 64(3):659-669.
- Popa KN, Chait LD, Johansson CE. Reinforcing and subjective effects of caffeine in normal human volunteers. Psychopharmacology 1989; 98:81–88.
- Evans SM, Griffiths RR. Dose-related caffeine discrimination in normal volunteers: individual differences in subjective and self-reported cues. Behav Pharmacol 1991; 2:345–356.
- Luciano M, Kirk KM, Heath AC, Martin NG. The genetics of tea and coffee drinking and preference for source of caffeine in a large community sample of Australian twins. Addiction. 2005; 100(10):1510-1517.
- Cornelis MC, El-Sohemy A, Campos H. Genetic polymorphism of the adenosine A2A receptor is associated with habitual caffeine consumption. Am J Clin Nutr 2007; 86(1):240-244.
- Childs E, Hohoff C, Deckert J, Xu K, Badner J, De Wit H. Association between ADORA2A and DRD2 Polymorphisms and Caffeine-Induced Anxiety. Neuropsychopharmacology 2008; 33:2791–2800.
- Rogers PJ, Hohoff C, Heatherley SV, Mullings EL, Maxfield PJ, Evershed RP, Deckert J, Nutt DJ.Association of the Anxiogenic and Alerting Effects of Caffeine with ADORA2A and ADORA1 Polymorphisms and Habitual Level of Caffeine Consumption. Neuropsychopharmacology 2010; 35:1973–1983.
- Riksen N P, Franke B, van den Broek P, Smits P, Rongen GA. The 1976C>T polymorphism in the adenosine A2A receptor gene does not affect the vasodilator response to adenosine in humans in vivo. Pharmacogenetics and Genomics 2007; 17(7):551-554.
- Rétey JV, Adam M, Khatami R,Luhmann UFO, Jung HH, Berger W, Landolt H-P. A Genetic Variation in the Adenosine A2A Receptor Gene (ADORA2A) Contributes to Individual Sensitivity to Caffeine Effects on Sleep. Clin Pharmacol Ther 2007; 81(5):692-698.
- Benowitz NL, Jacob P, Mayan H, Denaro C. Sympathomimetic effects of paraxanthine and caffeine in humans. Clin Pharmacol Ther. 1995; 58(6):684-91.
- Fisone G, Borgkvist A, Usiello A. Caffeine as a psychomotor stimulant: mechanism of action. Cell Mol Life Sci 2004; 61: 857-72.
- Daly JW, Jacobson KA, Ukena D. Adenosine receptors: development of selective agonists and antagonists. Prog Clin Biol Res. 1987; 230(1):41-63.
- Chang D, Song D, Zhang J, Shang Y, Ge Q, Wang Z . Caffeine Caused a Widespread Increase of Resting Brain Entropy. Sci Rep 2018; 8:2700-2707.
- Vidyasagar R, Greyling A, Draijer R, Corfield DR, Parkes LM. The effect of black tea and caffeine on regional cerebral blood flow measured with arterial spin labeling. J Cereb Blood Flow Metab. 2013; 33(6):963-8.
- Vandenberghe C, St-Pierre V, Courchesne-Loyer A, Hennebelle M, Castellano CA, Cunnane SC Caffeine intake increases plasma ketones: an acute metabolic study in humans. Can J Physiol Pharmacol. 2017; 95(4):455-458.
- Clark I, Landolt HP. Coffee, caffeine, and sleep: A systematic review of epidemiological studies and randomized controlled trials. Sleep Med Rev. 2017; 31:70–8.
- Metro D., Cernaro V., Santoro D., Papa M., Buemi M., Benvenga S., Manasseri L. Beneficial effects of oral pure caffeine on oxidative stress. J. Clin. Transl. Endocrinol. 2017;10:22–27.
- Walter S, Letiembre M, Liu Y, et al..Role of the toll-like receptor 4 in neuroinflammation in Alzheimer's disease. Int J Exp Cell Physiol Biochem Pharm, 2007; 20(6):947-956.
- Badshah H, Ikram M, Ali W, Ahmad A, Hahm JR, Kim MO. Caffeine May Abrogate LPS-Induced Oxidative Stress and Neuroinflammation by Regulating Nrf2/TLR4 in Adult Mouse Brains. Biomolecules. 2019; 9(11):719-731.
- Selkoe DJ, Hardy J. The amyloid hypothesis of alzheimer's disease at 25 years. EMBO Mol Med. 2016; 8(6): 595–608.
- Popugaeva E, Pchitskaya E, Bezprozvanny I. Dysregulation of neuronal calcium homeostasis in alzheimer's disease - a therapeutic opportunity? Biochem Biophys Res Commun 2017; 483(4):998–1004.
- Chui DH, Tanahashi H, Ozawa K, Ikeda S, Checler F, Ueda O. et al. Transgenic mice with alzheimer presenilin 1 mutations show accelerated neurodegeneration without amyloid plaque formation. Nat Med.1999; 5(5):560–4.
- Gomes CV, Kaster MP, Tome AR, Agostinho PM, Cunha RA. Adenosine receptors and brain diseases: Neuroprotection and neurodegeneration.Biochim Biophys Acta 2011; 1808(5):1380–99.
- Rozov A, Burnashev N. Fast interaction between ampa and nmda receptors by intracellular calcium. Cell Calcium 2016;60(6):407–14.
- Yamazawa T, Murayama T, Oyamada H, Suzuki J, Kurebayashi N, Kanemaru K. et al. Correlation of molecular dynamics analysis and calcium signaling in mutant ryanodine receptors. Biophys J 2016; 110(3):263a.
- Santulli G, Marks AR. Essential Roles of Intracellular Calcium Release Channels in Muscle, Brain, Metabolism, and Aging. Current Molecular Pharmacology, 2015; 8(2):206–22.
- Zucchi R, Ronca-Testoni S. The sarcoplasmic reticulum Ca2+ channel/ryanodine receptor: modulation by endogenous effectors, drugs and disease states. Pharmacological Reviews, 1997; 49(1):1–51.
- Chan S L, Mayne M, Holden C P, Geiger J D, Mattson M P. Presenilin-1 mutations increase levels of ryanodine receptors and calcium release in PC12 cells and cortical neurons. J Biol Chem 2000; 275(24):18195-200
- Danysz W, Parsons CG. Alzheimer's disease, beta-amyloid, glutamate, nmda receptors and memantine--searching for the connections. Br J Pharmacol 2012; 167(2):324–52.
- Del Prete D, Checler F, Mounia Chami M. Ryanodine receptors: physiological function and deregulation in Alzheimer disease. Mol Neurodegeneration 2014; 9:21-36.
- Lacampagne A, Liu X, Reiken S, Bussiere R, Meli AC, Lauritzen I, et al. Post-translational remodeling of ryanodine receptor induces calcium leak leading to alzheimer's disease-like pathologies and cognitive deficits. Acta Neuropathol 2017. Acta Neuropathologica 2017; 134(5):749-767.
- Bussiere R, Lacampagne A, Reiken S, Liu X, Scheuerman V, Zalk R, Martin C, Checler F, Marks AR, Chami M. Post-translational remodeling of ryanodine receptor induces calcium leak leading to Alzheimer's disease-like pathologies and cognitive deficits. J Biol Chem, 2017; 292(24):10153-10168.
- Pérez A, Li T, Hernandez S, Zhang R, Cao C. The rationale of using coffee and melatonin as an alternative treatment for alzheimer’s disease. J Alzheimer Dis Parkinsonism. 2016; 6: 205.
- Maia L, de Mendonca A. Does caffeine intake protect from alzheimer's disease? Eur J Neurol. 2002; 9(4):377–82.
- de Mendonça A, Sebastião AM, Ribeiro JA. Adenosine: Does it have a neuroprotective role after all? Brain Res Rev. 2000; 33(2):258–74.
- Keshavarz M, Farrokhi MR, Amiri A. Caffeine Neuroprotective Mechanism Against β-Amyloid Neurotoxicity in SHSY5Y Cell Line: Involvement of Adenosine, Ryanodine, and N-Methyl-D-Aspartate Receptors. Advan Pharm Bull, 2017; 7(4):579-584.
- Popat R A, Van Den Eeden S K, Tanner CM, et al.. Coffee, ADORA2A, and CYP1A2: the caffeine connection in Parkinson’s disease. Eur J Neurol 2011; 18(5): 759-765.
- Fujimaki M, Saiki S, Li Y, et al.. Serum caffeine and metabolites are reliable biomarkers of early Parkinson disease. Neurology 2018; 90(5):e1-e8.
- Deborah Brauser. Low Levels of Caffeine, Metabolites May Identify Early Parkinson's Disease -Medscape- 5 Jan 05, 2018.
- Crotty GF, Maciuca R, Macklin EA, et al. Association of caffeine and related analytes with resistance to Parkinson disease among LRRK2 mutation carriers. A metabolomic study. Neurology 2020; 95(24):e3428-e3437.
- Hong CT, Chan L, Bai C-H. The Effect of Caffeine on the Risk and Progression of Parkinson’s Disease: A Meta-Analysis. Nutrients 2020; 12(6), 1860.
- Altabakhi IW, Anderson J, Zito PM. Acetaminophen/Aspirin/Caffeine. In: StatPearls. Treasure Island (FL): StatPearls Publishing; September 29, 2020.
- Raskin NH. The hypnic Headache syndrome. Headache. 1988; 28:534-536.
- Pereira Silva-Néto R, Mesquita Sousa Santos PE , Prieto Peres MF. Hypnic headache: A review of 348 cases published from 1988 to 2018. J Neurol Sci. 2019; 401:103-109.
- Lipton RB, Diener HC , Robbins MS , Garas SY, Patel K. Caffeine in the management of patients with headache. J Headache Pain. 2017; 18(1):107-117.
- Grobbee DE, Rimm EB, Giovannucci E, Colditz G, Stampfer M, Willett W. Coffee, caffeine, and cardiovascular disease in men. N Engl J Med. 1990; 323:1026–1032.
- Hakim AA, Ross GW, Curb JD, Rodriguez BL, Burchfiel CM, Sharp DS, et al.. Coffee consumption in hypertensive men in older middle-age and the risk of stroke: the Honolulu Heart Program. J Clin Epidemiol. 1998; 51:487–494.
- Bidel S, Hu G, Qiao Q, Jousilahti P, Antikainen R, Tuomilehto J. Coffee consumption and risk of total and cardiovascular mortality among patients with type 2 diabetes. Diabetologia. 2006; 49:2618–2626.
- Larsson SC, Männistö S, Virtanen MJ, Kontto J, Albanes D, Virtamo J. Coffee and tea consumption and risk of stroke subtypes in male smokers. Stroke. 2008; 39:1681–1687.
- Lopez-Garcia E, Rodriguez-Artalejo F, Rexrode KM, Logroscino G, Hu FB, van Dam RM. Coffee consumption and risk of stroke in women. Circulation. 2009; 119:1116–1123.
- Lee SM, Choi NK, Lee BC, Cho KH, Yoon BW, Park BJ. Caffeine-containing medicines increase the risk of hemorrhagic stroke. Stroke. 2013; 44(8):2139-43.
- Saab S, Mallam D, Cox GA, Tong MJ. Impact of coffee on liver diseases: a systematic review. Liver Int, 2014; 34(4):495-504.
- Kennedy OJ, Roderick P, Buchanan R, et al.. Coffee, including caffeinated and decaffeinated coffee, and the risk of hepatocellular carcinoma: a systematic review and dose–response meta-analysis BMJ Open 2017; 7:e013739. doi: 10.1136/bmjopen-2016-013739.
- Bonita JS, Mandarano M, Shuta D, Vinson J. Coffee and cardiovascular disease: in vitro, cellular, animal, and human studies. Pharmacol Res. 2007; 55:187–198.
- Robertson D, Frölich JC, Carr RK, Watson JT, Hollifield JW, Shand DG, Oates JA. Effects of caffeine on plasma renin activity, catecholamines and blood pressure. N Engl J Med. 1978; 298:181–186.
- Wendt IR, Stephenson DG. Effects of caffeine on Ca‐activated force production in skinned cardiac and skeletal muscle fibres of the rat. Pflugers Arch. 1983; 398:210–216.
- Kong H, Jones PP, Koop A, Zhang L, Duff HJ, Wayne Chen SR. Caffeine induces Ca2+ release by reducing the threshold for luminal Ca2+ activation of the ryanodine receptor. Biochem J. 2008; 414:441–452.
- Conen D, Chiuve SE, Everett BM, Zhang SM, Buring JE, Albert CM. Caffeine consumption and incident atrial fibrillation in women. Am J Clin Nutr. 2010; 92:509–514.
- Larsson SC, Drca N, Jensen‐Urstad M, Wolk A. Coffee consumption is not associated with increased risk of atrial fibrillation: results from two prospective cohorts and a meta‐analysis. BMC Med. 2015; 13:207.
- Voskoboinik A, Koh Y, Kistler PM. Cardiovascular effects of caffeinated beverages. Trends Cardiovasc Med, 2019; 29(6):345-350.
- Bodar V, Chen J, Gaziano JM, Albert C, Djoussé L. Coffee Consumption and Risk of Atrial Fibrillation in the Physicians’ Health Study. J Am Heart Assoc 2019; 8(15):e011346.
- Kim EJ, Nah G, Vittinghoff E, Delling FB, Marcus GM. Is drinking coffe associated with an increased risk of arrhythmia? Heart Rhythm Society (HRS) 2020: Póster D-PO01-032.
- Vieira MB, Magriço R, Dias CV, Leitão L, Neves JS. Caffeine consumption and mortality in chronic kidney disease: a nationally representative analysis. Nephrol Dial Transplant 2019; 34(6):974–980.
No hay comentarios:
Publicar un comentario